Cyclin-abhängiger Kinase-Komplex

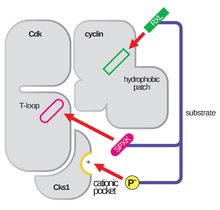
Die Struktur von CDKs im Komplex mit einer Cyclin-Untereinheit (CDKC) ist seit langem ein Ziel von Struktur- und Zellbiologen, beginnend in den 1990er Jahren, als die Struktur von ungebundenem Cyclin A von Brown et al. gelöst wurde und im selben Jahr Jeffery et al. die Struktur des menschlichen Cyclin A-CDK2-Komplexes mit einer Auflösung von 2,3 Angström auflösten. Seit dieser Zeit wurden viele CDK-Strukturen mit höherer Auflösung bestimmt, einschließlich der Strukturen von CDK2 und CDK2 gebunden an eine Vielzahl von Substraten, wie in Abbildung 1 zu sehen ist.Hochauflösende Strukturen existieren für insgesamt etwa 25 CDK-Cyclin-Komplexe in der Protein Data Bank. Basierend auf der Funktion gibt es zwei allgemeine Populationen von CDK-Cyclin-Komplexstrukturen, die offene und die geschlossene Form. Der Unterschied zwischen den Formen liegt in der Bindung der Cyclinpartner, wobei bei Komplexen der geschlossenen Form die CDK-Cyclin-Bindung sowohl am C- als auch am N-Terminus der Aktivierungsschleife der CDK erfolgt, während die Partner der offenen Form nur am N-Terminus binden. Strukturen der offenen Form entsprechen am häufigsten den Komplexen, die an der Transkriptionsregulation beteiligt sind (CDK 8, 9, 12 und 13), während CDK-Cyclin-Komplexe der geschlossenen Form am häufigsten an der Zellzyklusprogression und -regulation beteiligt sind (CDK 1, 2, 6). Diese unterschiedlichen Rollen unterscheiden sich jedoch nicht wesentlich durch die Sequenzhomologie zwischen den CDK-Komponenten. Insbesondere scheint es unter diesen bekannten Strukturen vier große konservierte Regionen zu geben: eine N-terminale Glycin-reiche Schleife, eine Hinge-Region, eine αC-Helix und eine T-Schleifen-Regulationsstelle.
Aktivierungsschleife
Die Aktivierungsschleife, auch als T-Schleife bezeichnet, ist die Region von CDK (zwischen den DFG- und APE-Motiven in vielen CDK), die enzymatisch aktiv ist, wenn CDK an seinen funktionsspezifischen Partner gebunden ist. In CDK-Cyclin-Komplexen besteht diese Aktivierungsregion aus einer konservierten αL-12-Helix und enthält einen phosphorylierbaren Schlüsselrest (normalerweise Threonin für CDK-Cyclin-Partner, aber auch Serin und Tyrosin), der die enzymatische Aktivität der CDK vermittelt. An diesem essentiellen Rest (T160 in CDK2-Komplexen, T177 in CDK6-Komplexen) findet die enzymatische ATP-Phosphorylierung von CDK-Cyclin-Komplexen durch CAK (cyclin activating kinase, bezogen auf den CDK7-Cyclin H-Komplex in menschlichen Zellen) statt. Nach der Hydrolyse von ATP zur Phosphorylierung an dieser Stelle sind diese Komplexe in der Lage, ihre vorgesehene Funktion, die Phosphorylierung von zellulären Zielen, zu erfüllen. Es ist wichtig zu beachten, dass bei CDK 1, 2 und 6 die T-Schleife und eine separate C-terminale Region die Hauptbindungsstellen für Cycline sind, und welche Cycline an jeden dieser CDKs gebunden werden, wird durch die besondere Sequenz der Aktivierungsstelle T-Schleife vermittelt. Diese Cyclin-Bindungsstellen sind die Regionen mit der höchsten Variabilität in CDKs, trotz der relativ hohen Sequenzhomologie, die das αL-12-Helix-Motiv dieser Strukturkomponente umgibt.
Glycin-reiche RegionBearbeiten
Die glycinreiche Schleife (Gly-rich loop), wie sie in den Resten 12-16 in CDK2 zu sehen ist, kodiert ein konserviertes GXGXXG-Motiv sowohl in Hefe als auch im Tiermodell. Die regulatorische Region unterliegt einer differentiellen Phosphorylierung an Nicht-Glycin-Resten innerhalb dieses Motivs, so dass diese Stelle in Säugetieren einer Wee1- und/oder Myt1-inhibitorischen Kinase-Phosphorylierung und Cdc25-Dephosphorylierung unterliegt. Diese reversible Phosphorylierung an der Gly-reichen Schleife in CDK2 erfolgt an Y15, wo die Aktivität weiter untersucht wurde. Die Untersuchung dieses Restes hat gezeigt, dass die Phosphorylierung eine Konformationsänderung fördert, die die ATP- und Substratbindung durch sterische Interferenz mit diesen notwendigen Bindungsstellen in der Aktivierungsschleife der CDK-Cyclin-Komplexe verhindert. Diese Aktivität wird durch die bemerkenswerte Flexibilität unterstützt, die die Gly-reiche Schleife innerhalb der Struktur der meisten CDK hat, so dass ihre Rotation in Richtung der Aktivierungsschleife einen signifikanten Effekt auf die Verringerung der Substrat-Affinität hat, ohne größere Veränderungen in der Gesamtstruktur des CDK-Cyclin-Komplexes.
Scharnierregion
Die konservierte Scharnierregion von CDK in eukaryotischen Zellen fungiert als eine wesentliche Brücke zwischen der Gly-reichen Schleife und der Aktivierungsschleife. CDK sind durch einen N-terminalen Lappen charakterisiert, der hauptsächlich aus einem verdrillten Beta-Faltblatt besteht und über diese Scharnierregion mit einem von einer Alpha-Helix dominierten C-terminalen Lappen verbunden ist. Bei der Diskussion der T-Schleife und der Gly-reichen Schleife ist es wichtig zu beachten, dass diese Regionen, die in der Lage sein müssen, räumlich zu interagieren, um ihre biochemischen Funktionen auszuführen, auf gegenüberliegenden Lappen der CDK selbst liegen. Somit verbindet diese Scharnierregion, deren Länge je nach CDK-Typ und CDK-Cyclin-Komplex leicht variieren kann, wesentliche regulatorische Regionen der CDK, indem sie diese Lappen miteinander verbindet, und spielt eine Schlüsselrolle in der resultierenden Struktur von CDK-Cyclin-Komplexen, indem sie ATP richtig ausrichtet, um die Katalyse von Phosphorylierungsreaktionen durch den zusammengesetzten Komplex zu erleichtern.
αC-HelixEdit
Die αC-Helix-Region ist in vielen Bereichen des Säugetierkinoms (Familie der Kinasen) hoch konserviert. Ihre Hauptaufgabe ist es, die allosterische Kontrolle über das aktive Zentrum der Kinase aufrechtzuerhalten. Diese Kontrolle manifestiert sich in CDK-Cyclin-Komplexen, indem die CDK-Aktivität spezifisch verhindert wird, bis sie an ihren Partnerregulator (d.h. Cyclin oder ein anderes Partnerprotein) bindet. Diese Bindung bewirkt eine Konformationsänderung in der αC-Helix-Region der CDK und ermöglicht es ihr, aus dem Spalt des aktiven Zentrums bewegt zu werden und den anfänglichen Prozess der T-Loop-Aktivierung abzuschließen. Da diese Region in der gesamten Protein-Superfamilie der Kinasen so konserviert ist, wird dieser Mechanismus, bei dem die αC-Helix nachweislich aus dem N-terminalen Lappen der Kinase herausgefaltet wird, was einen verbesserten Zugang zur αL-12-Helix ermöglicht, die innerhalb der T-Schleife liegt, als potenzielles Ziel für die Entwicklung von Medikamenten angesehen.