Complexo cinase dependente de ciclina

LoopEdit de Activação
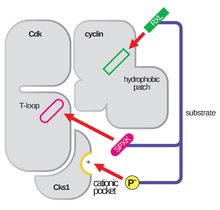
O laço de activação, também referido como o laço T, é a região do CDK (entre os motivos DFG e APE em muitos CDK) que é enzimaticamente activa quando o CDK está ligado ao seu parceiro específico de função. Nos complexos CDK-cyclin, esta região de activação é composta por um Helix conservado αL-12 e contém um resíduo fosforilatável chave (geralmente Threonine para parceiros CDK-cyclin, mas também inclui Serine e Tyrosine) que medeia a actividade enzimática do CDK. É neste resíduo essencial (T160 nos complexos CDK2, T177 nos complexos CDK6) que ocorre a fosforilação enzimática ATP dos complexos CDK-cyclin por CAK (ciclina activando a cinase, referindo-se ao complexo CDK7-Cyclin H em células humanas). Após a hidrólise de ATP para fosforilato neste local, estes complexos são capazes de completar a sua função pretendida, a fosforilação de alvos celulares. É importante notar que no CDK 1, 2 e 6, o T-loop e uma região C-terminal separada são os principais locais de ligação da ciclina no CDK, e quais as ciclinas ligadas a cada um destes CDK são mediadas pela sequência particular do T-loop do local de activação. Estes locais de ligação da ciclina são as regiões de maior variabilidade no CDK, apesar da homologia relativamente elevada da sequência homóloga em torno do motivo Helix αL-12 deste componente estrutural.
Região rica em glicinaEdit
O loop rico em glicina (loop rico em glicina) como se vê nos resíduos 12-16 no CDK2 codifica um motivo GXGXXG conservado em ambos os modelos de leveduras e animais. A região reguladora está sujeita à fosforilação diferencial a resíduos não glicosilados dentro deste motivo, tornando este site sujeito à fosforilação inibitória da cinase Wee1 e/ou Myt1 e à desfosforilação do Cdc25 em mamíferos. Esta fosforilação reversível no laço rico em glioxina no CDK2 ocorre no Y15, onde a actividade tem sido mais estudada. O estudo deste resíduo demonstrou que a fosforilação promove uma alteração conformacional que impede a ligação do ATP e do substrato por interferência estérica com estes locais de ligação necessários no laço de activação dos complexos CDK-cyclin. Esta actividade é auxiliada pela notável flexibilidade que o laço rico em glândulas tem dentro da estrutura da maioria dos CDK, permitindo que a sua rotação em direcção ao laço de activação tenha um efeito significativo na redução da afinidade do substrato sem grandes alterações na estrutura global do complexo CDK-cyclin.
Região da dobradiçaEditar
A região conservada da dobradiça CDK dentro das células eucarióticas actua como uma ponte essencial entre o laço rico em glândulas e o laço de activação. Os CDK caracterizam-se por um lóbulo terminal N que é principalmente uma folha beta torcida ligada através desta região da dobradiça a um lóbulo terminal C dominado por uma hélice alfa. Na discussão do lóbulo T e do laço rico em Gly, é importante notar que estas regiões, que devem ser capazes de interagir espacialmente a fim de desempenharem as suas funções bioquímicas, encontram-se em lóbulos opostos do próprio CDK. Assim, esta região de dobradiça, que pode variar ligeiramente em comprimento entre o tipo CDK e o complexo CDK-cyclin, liga regiões reguladoras essenciais do CDK ligando estes lóbulos, e desempenha papéis-chave na estrutura resultante dos complexos CDK-cyclin, orientando adequadamente o ATP para uma fácil catálise das reacções de fosforilação pelo complexo montado.
αC-HelixEdit
A região αC-Helix está altamente conservada através de muitos dos kinomas de mamíferos (família das kinases). A sua principal responsabilidade é manter o controlo alérgico do sítio activo do kinase. Este controlo manifesta-se em complexos de CDK-cyclin, impedindo especificamente a actividade CDK até à sua ligação ao seu regulador parceiro (ou seja, ciclina ou outra proteína parceira). Esta ligação provoca uma mudança conformacional na região de αC-Helix do CDK e permite que este seja deslocado da fenda do sítio activo e completa o processo inicial de activação do T-loop. Dado que esta região está tão conservada através da superfamília proteica das kinases, este mecanismo onde o αC-Helix demonstrou dobrar-se para fora do lóbulo N-terminal da kinase, permitindo um maior acesso à Helix αL-12 que se encontra dentro do laço T, é considerado um alvo potencial para o desenvolvimento de fármacos.