Complexe kinase dépendant des cyclines

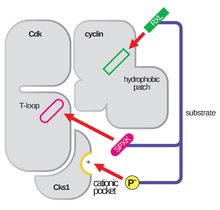
La structure des CDK en complexe avec une sous-unité de cycline (CDKC) est depuis longtemps un objectif des biologistes structuraux et cellulaires, à partir des années 1990 lorsque la structure de la cycline A non liée a été résolue par Brown et al. et la même année, Jeffery et al. ont résolu la structure du complexe cycline A-CDK2 humain à une résolution de 2,3 Angstrom. Depuis, de nombreuses structures de CDK ont été déterminées à une plus haute résolution, y compris les structures de CDK2 et de CDK2 liée à une variété de substrats, comme on peut le voir sur la figure 1.Des structures à haute résolution existent pour environ 25 complexes CDK-cycline au total dans la banque de données des protéines. Sur la base de la fonction, il existe deux populations générales de structures de complexes CDK-cycline, la forme ouverte et la forme fermée. La différence entre les formes réside dans la liaison des partenaires de la cycline, où les complexes de forme fermée ont une liaison CDK-cycline à la fois à l’extrémité C et à l’extrémité N de la boucle d’activation de la CDK, tandis que les partenaires de forme ouverte se lient uniquement à l’extrémité N. Les structures de forme ouverte correspondent le plus souvent aux complexes impliqués dans la régulation transcriptionnelle (CDK 8, 9, 12 et 13), tandis que les complexes CDK-cycline de forme fermée sont le plus souvent impliqués dans la progression et la régulation du cycle cellulaire (CDK 1, 2, 6). Ces rôles distincts, cependant, ne diffèrent pas de manière significative avec l’homologie de séquence entre les composants CDK. En particulier, parmi ces structures connues, il semble y avoir quatre régions conservées majeures : une boucle N-terminale riche en glycine, une région charnière, une hélice αC et un site de régulation de la boucle en T.
La boucle d’activation, également appelée boucle en T, est la région de la CDK (entre les motifs DFG et APE dans de nombreuses CDK) qui est enzymatiquement active lorsque la CDK est liée à son partenaire spécifique à la fonction. Dans les complexes CDK-cycline, cette région d’activation est composée d’une hélice αL-12 conservée et contient un résidu phosphorylable clé (généralement la thréonine pour les partenaires CDK-cycline, mais aussi la sérine et la tyrosine) qui intervient dans l’activité enzymatique de la CDK. C’est au niveau de ce résidu essentiel (T160 dans les complexes CDK2, T177 dans les complexes CDK6) qu’a lieu la phosphorylation enzymatique par l’ATP des complexes CDK-cycline par la CAK (cyclin activating kinase, faisant référence au complexe CDK7-Cyclin H dans les cellules humaines). Après l’hydrolyse de l’ATP pour la phosphorylation à ce site, ces complexes sont en mesure d’accomplir leur fonction prévue, la phosphorylation des cibles cellulaires. Il est important de noter que dans les CDK 1, 2 et 6, la boucle en T et une région C-terminale distincte sont les principaux sites de liaison des cyclines dans la CDK, et que les cyclines qui sont liées à chacune de ces CDK sont médiées par la séquence particulière de la boucle en T du site d’activation. Ces sites de liaison des cyclines sont les régions de plus grande variabilité dans les CDK malgré une homologie de séquence relativement élevée entourant le motif αL-12 Helix de ce composant structurel.
Région riche en glycineEdit
La boucle riche en glycine (boucle riche en Gly) telle que vue dans les résidus 12-16 dans CDK2 code un motif GXGXXG conservé à travers la levure et les modèles animaux. La région régulatrice est soumise à une phosphorylation différentielle au niveau des résidus non glycine au sein de ce motif, ce qui fait que ce site est soumis à la phosphorylation des kinases inhibitrices Wee1 et/ou Myt1 et à la dé-phosphorylation de Cdc25 chez les mammifères. Cette phosphorylation réversible au niveau de la boucle riche en Gly dans CDK2 se produit à Y15, dont l’activité a été étudiée plus en détail. L’étude de ce résidu a montré que la phosphorylation favorise un changement de conformation qui empêche la liaison de l’ATP et du substrat par interférence stérique avec ces sites de liaison nécessaires dans la boucle d’activation des complexes CDK-cycline. Cette activité est favorisée par la flexibilité notable que possède la boucle riche en Gly dans la structure de la plupart des CDK permettant à sa rotation vers la boucle d’activation d’avoir un effet significatif sur la réduction de l’affinité du substrat sans changement majeur dans la structure globale du complexe CDK-cycline.
Région charnièreEdit
La région charnière conservée des CDK au sein des cellules eucaryotes agit comme un pont essentiel entre la boucle riche en Gly et la boucle d’activation. Les CDK sont caractérisées par un lobe N-terminal qui est principalement un feuillet bêta torsadé connecté via cette région charnière à un lobe C-terminal dominé par une hélice alpha. Dans la discussion sur la boucle T et la boucle riche en Gly, il est important de noter que ces régions, qui doivent pouvoir interagir dans l’espace afin de remplir leurs fonctions biochimiques, se trouvent sur des lobes opposés de la CDK elle-même. Ainsi, cette région charnière, dont la longueur peut varier légèrement entre le type de CDK et le complexe CDK-cycline, relie des régions régulatrices essentielles de la CDK en connectant ces lobes, et joue des rôles clés dans la structure résultante des complexes CDK-cycline en orientant correctement l’ATP pour faciliter la catalyse des réactions de phosphorylation par le complexe assemblé.
Edition αC-Helix
La région αC-Helix est hautement conservée à travers de nombreux kinomes (famille des kinases) de mammifères. Sa principale responsabilité est de maintenir le contrôle allostérique du site actif de la kinase. Ce contrôle se manifeste dans les complexes CDK-cycline en empêchant spécifiquement l’activité de la CDK jusqu’à ce qu’elle se lie à son partenaire régulateur (c’est-à-dire la cycline ou une autre protéine partenaire). Cette liaison provoque un changement de conformation de la région αC-Helix de la CDK et lui permet d’être déplacée de la fente du site actif et de compléter le processus initial d’activation de la boucle T. Étant donné que cette région est tellement conservée dans la superfamille des protéines kinases, ce mécanisme où il a été démontré que l’αC-Helix se replie hors du lobe N-terminal de la kinase, permettant un accès accru à l’αL-12 Helix qui se trouve dans la boucle en T, est considéré comme une cible potentielle pour le développement de médicaments.