Kompleks kinazy zależnej od cykliny

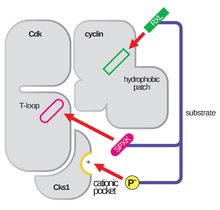
Struktura CDK w kompleksie z podjednostkami cykliny (CDKC) od dawna była celem biologów strukturalnych i komórkowych, począwszy od lat 90. XX wieku, kiedy to struktura niezwiązanej cykliny A została rozwiązana przez Browna i wsp. i w tym samym roku Jeffery i wsp. rozwiązali strukturę kompleksu ludzkiej cykliny A-CDK2 w rozdzielczości 2,3 Angstroma. Od tego czasu wiele struktur CDK zostało oznaczonych w wyższej rozdzielczości, w tym struktury CDK2 i CDK2 związanej z różnymi substratami, jak widać na Rysunku 1. W Protein Data Bank istnieją struktury o wysokiej rozdzielczości dla około 25 kompleksów CDK-cyklina. W oparciu o funkcję, istnieją dwie ogólne populacje struktur kompleksu CDK-cyklina, forma otwarta i zamknięta. Różnica pomiędzy tymi formami polega na wiązaniu partnerów cyklinowych, gdzie kompleksy w formie zamkniętej wiążą CDK-cykliny zarówno na C jak i N-końcu pętli aktywacyjnej CDK, podczas gdy partnerzy w formie otwartej wiążą się tylko na N-końcu. Struktury w formie otwartej odpowiadają najczęściej kompleksom zaangażowanym w regulację transkrypcji (CDK 8, 9, 12 i 13), podczas gdy kompleksy CDK-cyklina w formie zamkniętej są najczęściej zaangażowane w progresję i regulację cyklu komórkowego (CDK 1, 2, 6). Te odrębne role nie różnią się jednak znacząco od homologii sekwencji pomiędzy składnikami CDK. W szczególności, wśród tych znanych struktur wydają się być cztery główne konserwowane regiony: N-końcowa pętla bogata w glicynę, region zawiasowy, helisa αC i miejsce regulacji pętli T.
Pętla aktywacyjnaEdit
Pętla aktywacyjna, zwana również pętlą T, jest regionem CDK (pomiędzy motywami DFG i APE w wielu CDK), który jest enzymatycznie aktywny, gdy CDK jest związana ze swoim specyficznym dla funkcji partnerem. W kompleksach CDK-cyklina ten region aktywacji składa się z konserwowanej helisy αL-12 i zawiera kluczową resztę fosforylowaną (zwykle treoninę w przypadku partnerów CDK-cyklina, ale także serynę i tyrozynę), która pośredniczy w aktywności enzymatycznej CDK. To właśnie przy tej niezbędnej reszcie (T160 w kompleksach CDK2, T177 w kompleksach CDK6) odbywa się enzymatyczna fosforylacja ATP kompleksów CDK-cyklina przez CAK (cyclin activating kinase, odnosząca się do kompleksu CDK7-Cyklina H w komórkach ludzkich). Po hydrolizie ATP do fosforylacji w tym miejscu, kompleksy te są w stanie dokończyć swoją zamierzoną funkcję, czyli fosforylację celów komórkowych. Ważne jest, aby zauważyć, że w CDK 1, 2 i 6, pętla T i oddzielny region C-końcowy są głównymi miejscami wiązania cykliny w CDK, a które cykliny są związane z każdym z tych CDK jest pośredniczony przez szczególną sekwencję pętli T miejsca aktywacji. Te miejsca wiązania cyklin są regionami o najwyższej zmienności w CDK, pomimo stosunkowo wysokiej homologii sekwencji otaczającej motyw αL-12 Helix tego elementu strukturalnego.
Region bogaty w glicynęEdit
Pętla bogata w glicynę (pętla bogata w glicynę) widoczna w resztach 12-16 w CDK2 koduje konserwowany motyw GXGXXG zarówno w modelach drożdżowych, jak i zwierzęcych. Region regulacyjny podlega zróżnicowanej fosforylacji przy resztach nieglicyny w obrębie tego motywu, co sprawia, że miejsce to podlega fosforylacji kinazy hamującej Wee1 i/lub Myt1 oraz de-fosforylacji Cdc25 u ssaków. Ta odwracalna fosforylacja w pętli bogatej w Gly w CDK2 występuje przy Y15, gdzie aktywność została dokładniej zbadana. Badania tej reszty wykazały, że fosforylacja promuje zmianę konformacyjną, która uniemożliwia wiązanie ATP i substratów poprzez steryczną ingerencję w te niezbędne miejsca wiązania w pętli aktywacyjnej kompleksów CDK-cyklina. Działanie to jest wspomagane przez znaczną elastyczność pętli bogatej w glicynę w strukturze większości CDK, co pozwala na jej rotację w kierunku pętli aktywacyjnej, co ma znaczący wpływ na zmniejszenie powinowactwa do substratu bez większych zmian w ogólnej strukturze kompleksu CDK-cyklina.
Region zawiasowyEdit
Zachowany region zawiasowy CDK w komórkach eukariotycznych działa jako istotny pomost pomiędzy pętlą bogatą w glicynę a pętlą aktywacyjną. CDK charakteryzują się N-końcowym płatem, który jest głównie skręconym arkuszem beta połączonym poprzez ten region zawiasowy z płatem C-końcowym zdominowanym przez helisę alfa. W dyskusji na temat pętli T i pętli bogatej w Gly należy zauważyć, że regiony te, które muszą być zdolne do przestrzennej interakcji, aby pełnić swoje funkcje biochemiczne, leżą na przeciwległych płatach samej CDK. Tak więc, ten region zawiasowy, którego długość może się nieznacznie różnić w zależności od typu CDK i kompleksu CDK-cyklina, łączy istotne regiony regulacyjne CDK poprzez połączenie tych płatów i odgrywa kluczową rolę w strukturze wynikowej kompleksów CDK-cyklina poprzez odpowiednie ukierunkowanie ATP w celu ułatwienia katalizy reakcji fosforylacji przez złożony kompleks.
αC-HelixEdit
Region αC-Helix jest wysoce konserwowany w wielu kinomach ssaków (rodzina kinaz). Jego głównym zadaniem jest utrzymanie kontroli allosterycznej nad miejscem aktywnym kinazy. Kontrola ta przejawia się w kompleksach CDK-cyklina poprzez specyficzne zapobieganie aktywności CDK do momentu związania się z jej partnerem regulatorowym (tj. cykliną lub innym białkiem partnerskim). To wi±zanie powoduje zmianę konformacyjn± w regionie αC-Helix CDK, co pozwala na jego przemieszczenie z miejsca aktywnego i zakończenie procesu aktywacji pętli T. Biorąc pod uwagę fakt, że region ten jest tak dobrze zachowany w całej rodzinie kinaz białkowych, mechanizm, w którym αC-Helix jest składany z N-końcowego płata kinazy, umożliwiając zwiększony dostęp do αL-12 Helix, który leży w obrębie pętli T, jest uważany za potencjalny cel dla rozwoju leków.