Complejo de cinasas dependientes de ciclinas

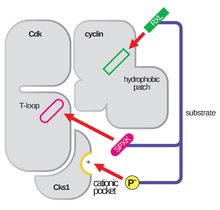
La estructura de las CDKs en complejo con una subunidad ciclina (CDKC) ha sido durante mucho tiempo un objetivo de los biólogos estructurales y celulares, comenzando en la década de 1990 cuando la estructura de la ciclina A no unida fue resuelta por Brown et al. y en el mismo año Jeffery et al. resolvieron la estructura del complejo ciclina A-CDK2 humana a una resolución de 2,3 Angstrom. Desde entonces, se han determinado muchas estructuras de CDK a mayor resolución, incluyendo las estructuras de CDK2 y CDK2 unida a una variedad de sustratos, como se ve en la Figura 1. Existen estructuras de alta resolución para aproximadamente 25 complejos CDK-ciclina en total dentro del Banco de Datos de Proteínas. Basándose en la función, hay dos poblaciones generales de estructuras de complejos CDK-ciclina, la forma abierta y la cerrada. La diferencia entre las formas radica en la unión de los socios de la ciclina, donde los complejos de forma cerrada tienen la unión CDK-ciclina tanto en el C como en el N-terminal del bucle de activación de la CDK, mientras que los socios de forma abierta se unen sólo en el N-terminal. Las estructuras de forma abierta corresponden con mayor frecuencia a los complejos implicados en la regulación transcripcional (CDK 8, 9, 12 y 13), mientras que los complejos CDK-ciclina de forma cerrada participan con mayor frecuencia en la progresión y regulación del ciclo celular (CDK 1, 2 y 6). Sin embargo, estas distintas funciones no difieren significativamente con la homología de secuencia entre los componentes de las CDK. En particular, entre estas estructuras conocidas parece haber cuatro regiones principales conservadas: un bucle rico en glicina N-terminal, una región de bisagra, una hélice αC y un sitio de regulación del bucle T.
Bucle de activaciónEditar
El bucle de activación, también denominado bucle T, es la región de la CDK (entre los motivos DFG y APE en muchas CDK) que es enzimáticamente activa cuando la CDK está unida a su pareja de función específica. En los complejos CDK-ciclina, esta región de activación se compone de una hélice αL-12 conservada y contiene un residuo clave fosforilable (normalmente treonina para los socios CDK-ciclina, pero también incluye serina y tirosina) que media la actividad enzimática de la CDK. Es en este residuo esencial (T160 en los complejos CDK2, T177 en los complejos CDK6) donde tiene lugar la fosforilación enzimática por ATP de los complejos CDK-ciclina por parte de la CAK (quinasa activadora de ciclinas, en referencia al complejo CDK7-Ciclina H en las células humanas). Tras la hidrólisis del ATP para fosforilar en este sitio, estos complejos son capaces de completar su función prevista, la fosforilación de objetivos celulares. Es importante señalar que en las CDK 1, 2 y 6, el bucle T y una región C-terminal separada son los principales sitios de unión de ciclinas en las CDK, y qué ciclinas se unen a cada una de estas CDK está mediado por la secuencia particular del bucle T del sitio de activación. Estos sitios de unión de ciclinas son las regiones de mayor variabilidad en las CDKs a pesar de la relativamente alta homología de secuencia que rodea el motivo αL-12 Helix de este componente estructural.
Región rica en glicinaEditar
El bucle rico en glicina (bucle rico en Gly) como se ve en los residuos 12-16 en CDK2 codifica un motivo GXGXXG conservado a través de modelos de levadura y animales. La región reguladora está sujeta a una fosforilación diferencial en los residuos que no son de glicina dentro de este motivo, lo que hace que este sitio esté sujeto a la fosforilación de la cinasa inhibidora Wee1 y/o Myt1 y a la desfosforilación de Cdc25 en los mamíferos. Esta fosforilación reversible en el bucle rico en Gly en CDK2 se produce en Y15, donde se ha estudiado más a fondo la actividad. El estudio de este residuo ha demostrado que la fosforilación promueve un cambio conformacional que impide la unión del ATP y del sustrato por interferencia estérica con estos sitios de unión necesarios en el bucle de activación de los complejos CDK-ciclina. Esta actividad se ve favorecida por la notable flexibilidad que tiene el bucle rico en glúcidos dentro de la estructura de la mayoría de las CDK, lo que permite que su rotación hacia el bucle de activación tenga un efecto significativo en la reducción de la afinidad por el sustrato sin que se produzcan cambios importantes en la estructura general del complejo CDK-ciclina.
Región bisagraEditar
La región bisagra conservada de las CDK dentro de las células eucariotas actúa como un puente esencial entre el bucle rico en glúcidos y el bucle de activación. Las CDK se caracterizan por un lóbulo N-terminal que es principalmente una hoja beta retorcida conectada a través de esta región bisagra a un lóbulo C-terminal dominado por hélices alfa. Al hablar del bucle T y del bucle rico en Gly, es importante señalar que estas regiones, que deben poder interactuar espacialmente para llevar a cabo sus funciones bioquímicas, se encuentran en lóbulos opuestos de la propia CDK. Así, esta región bisagra, que puede variar ligeramente en longitud entre el tipo de CDK y el complejo CDK-ciclina, conecta regiones reguladoras esenciales de la CDK conectando estos lóbulos, y desempeña funciones clave en la estructura resultante de los complejos CDK-ciclina al orientar adecuadamente el ATP para facilitar la catálisis de las reacciones de fosforilación por parte del complejo ensamblado.
αC-HelixEdit
La región αC-Helix está altamente conservada a través de muchos de los mamíferos del cinoma (familia de quinasas). Su principal responsabilidad es mantener el control alostérico del sitio activo de la quinasa. Este control se manifiesta en los complejos CDK-ciclina impidiendo específicamente la actividad de la CDK hasta que se une a su regulador asociado (es decir, la ciclina u otra proteína asociada). Esta unión provoca un cambio conformacional en la región αC-Helix de la CDK y permite que se desplace de la hendidura del sitio activo y complete el proceso inicial de activación del bucle T. Dado que esta región está tan conservada en toda la superfamilia de proteínas quinasas, este mecanismo en el que se ha demostrado que la αC-Hélice se pliega fuera del lóbulo N-terminal de la quinasa, permitiendo un mayor acceso a la αL-12 Hélice que se encuentra dentro del bucle T, se considera un objetivo potencial para el desarrollo de fármacos.