Complesso della chinasi ciclina-dipendente

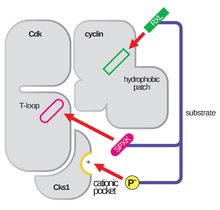
La struttura delle CDK in complesso con una subunità ciclina (CDKC) è stata a lungo un obiettivo dei biologi strutturali e cellulari a partire dagli anni ’90 quando la struttura della ciclina A non legata è stata risolta da Brown et al. e nello stesso anno Jeffery et al. hanno risolto la struttura del complesso ciclina A-CDK2 umana con una risoluzione di 2,3 Angstrom. Da allora, molte strutture di CDK sono state determinate ad alta risoluzione, comprese le strutture di CDK2 e CDK2 legate a una varietà di substrati, come si vede nella Figura 1. Esistono strutture ad alta risoluzione per circa 25 complessi CDK-ciclina in totale all’interno della Protein Data Bank. In base alla funzione, ci sono due popolazioni generali di strutture di complessi CDK-ciclina, forma aperta e chiusa. La differenza tra le forme risiede nel legame dei partner della ciclina, dove i complessi in forma chiusa hanno il legame CDK-ciclina sia al C che all’N-termine del loop di attivazione della CDK, mentre i partner in forma aperta si legano solo all’N-terminale. Le strutture in forma aperta corrispondono più spesso a quei complessi coinvolti nella regolazione trascrizionale (CDK 8, 9, 12, e 13), mentre i complessi CDK-ciclina in forma chiusa sono più spesso coinvolti nella progressione e regolazione del ciclo cellulare (CDK 1, 2, 6). Questi ruoli distinti, tuttavia, non differiscono significativamente con l’omologia di sequenza tra i componenti CDK. In particolare, tra queste strutture conosciute sembrano esserci quattro principali regioni conservate: un loop N-terminale ricco di glicina, una regione cerniera, un’αC-elica e un sito di regolazione T-loop.
Activation LoopEdit
Il loop di attivazione, chiamato anche T-loop, è la regione di CDK (tra i motivi DFG e APE in molte CDK) che è enzimaticamente attiva quando CDK è legata al suo partner specifico della funzione. Nei complessi CDK-ciclina, questa regione di attivazione è composta da un’elica conservata αL-12 e contiene un residuo chiave fosforilabile (di solito treonina per i partner CDK-ciclina, ma include anche serina e tirosina) che media l’attività enzimatica della CDK. È a questo residuo essenziale (T160 nei complessi CDK2, T177 nei complessi CDK6) che avviene la fosforilazione enzimatica dell’ATP dei complessi CDK-ciclina da parte della CAK (chinasi attivante la ciclina, riferita al complesso CDK7-Cyclin H nelle cellule umane). Dopo l’idrolisi dell’ATP per fosforilare in questo sito, questi complessi sono in grado di completare la loro funzione prevista, la fosforilazione dei bersagli cellulari. È importante notare che nelle CDK 1, 2 e 6, il T-loop e una regione C-terminale separata sono i principali siti di legame delle cicline nella CDK, e quali cicline sono legate a ciascuna di queste CDK è mediato dalla particolare sequenza del T-loop del sito di attivazione. Questi siti di legame delle cicline sono le regioni di maggiore variabilità nelle CDK nonostante l’omologia di sequenza relativamente alta che circonda il motivo αL-12 Helix di questo componente strutturale.
Regione ricca di glicinaModifica
L’anello ricco di glicina (Gly-rich loop) come visto nei residui 12-16 in CDK2 codifica un motivo GXGXXG conservato sia nel lievito che nei modelli animali. La regione regolatoria è soggetta a fosforilazione differenziale ai residui non glicinici all’interno di questo motivo, rendendo questo sito soggetto alla fosforilazione della chinasi inibitoria Wee1 e/o Myt1 e alla de-fosforilazione di Cdc25 nei mammiferi. Questa fosforilazione reversibile al loop ricco di Gly in CDK2 avviene a Y15, dove l’attività è stata ulteriormente studiata. Lo studio di questo residuo ha dimostrato che la fosforilazione promuove un cambiamento conformazionale che impedisce il legame dell’ATP e del substrato attraverso l’interferenza sterica con questi siti di legame necessari nell’anello di attivazione dei complessi CDK-ciclina. Questa attività è aiutata dalla notevole flessibilità che l’anello ricco di Gly ha all’interno della struttura della maggior parte delle CDK, permettendo la sua rotazione verso l’anello di attivazione per avere un effetto significativo sulla riduzione dell’affinità del substrato senza grandi cambiamenti nella struttura complessiva del complesso CDK-ciclina.
Regione della cernieraModifica
La regione della cerniera conservata della CDK all’interno delle cellule eucariotiche agisce come un ponte essenziale tra l’anello ricco di Gly e l’anello di attivazione. Le CDK sono caratterizzate da un lobo N-terminale che è principalmente un foglio beta attorcigliato collegato tramite questa regione di cerniera ad un lobo C-terminale dominato dall’alfa-elica. Nella discussione del T-loop e del Gly-rich loop, è importante notare che queste regioni, che devono essere in grado di interagire spazialmente per svolgere le loro funzioni biochimiche, si trovano su lobi opposti della CDK stessa. Quindi, questa regione cerniera, che può variare leggermente in lunghezza tra il tipo di CDK e il complesso CDK-ciclina, collega regioni regolatrici essenziali della CDK collegando questi lobi, e gioca ruoli chiave nella struttura risultante dei complessi CDK-ciclina orientando correttamente l’ATP per una facile catalisi delle reazioni di fosforilazione da parte del complesso assemblato.
αC-HelixEdit
La regione αC-Helix è altamente conservata in molte delle chinasi dei mammiferi. La sua responsabilità principale è quella di mantenere il controllo allosterico del sito attivo della chinasi. Questo controllo si manifesta nei complessi CDK-ciclina impedendo specificamente l’attività della CDK finché non si lega al suo regolatore partner (cioè la ciclina o un’altra proteina partner). Questo legame causa un cambiamento conformazionale nella regione αC-Helix della CDK e permette di spostarla dalla fessura del sito attivo e completa il processo iniziale di attivazione del T-loop. Dato che questa regione è così conservata in tutta la superfamiglia proteica delle chinasi, questo meccanismo in cui l’αC-Helix ha dimostrato di ripiegarsi fuori dal lobo N-terminale della chinasi, permettendo un maggiore accesso all’αL-12 Helix che si trova all’interno del T-loop, è considerato un potenziale bersaglio per lo sviluppo di farmaci.